Introduction
Les thymocytes sont les lymphocytes T immatures. Ils passent par différents stades caractérisés notamment par l’expression des molécules CD4 et CD8, d’où l’appellation des différents progéniteurs qui sont successivement double négatifs (CD4- CD8-), double positifs (CD4+ et CD8+) puis simple positifs (CD4+ ou CD8+).
Acquisition d’un TCR par recombinaison somatique
Étapes de sélection clonale
Activation spécifique suite à la rencontre antigénique
Pour simplifier le propos, seule l’ontogenèse des lymphocytes T αβ sera décrite ci-dessous.
Il existe des lymphocytes T γδ qui présentent un TCR γδ. Ils représentent environ 10 % du contingent lymphocytaire T et sont présents principalement dans les muqueuses. Certains d’entre eux peuvent reconnaitre des antigènes enchâssés dans des molécules HLA de classe I ou de classe II, néanmoins la plupart reconnaissent des ligands moins spécifiques :
- soit des lipides ou glycolipides enchâssés dans des molécules HLA non-classiques
- soit des protéines de choc thermique issue du soi
- soit des protéines d’origine microbienne
Acquisition d’un TCR par recombinaison somatique
Les caractéristiques structurelles du TCR ainsi que la recombinaison V(D)J du TCR sont décrites ici.
Où ?
Dans le thymus, dans la zone corticale (sous-capsulaire).
Quand ?
Au stade précoce du développement du thymocyte, lorsqu’il est double négatif (CD4- CD8-).
Mecanisme ?
Le réarrangement DJ, puis VDJ de la chaine β intervient en premier. Une fois la chaine β synthétisée, elle s’associe avec la chaine invariante pré-Tα pour constituer le pré-TCR.
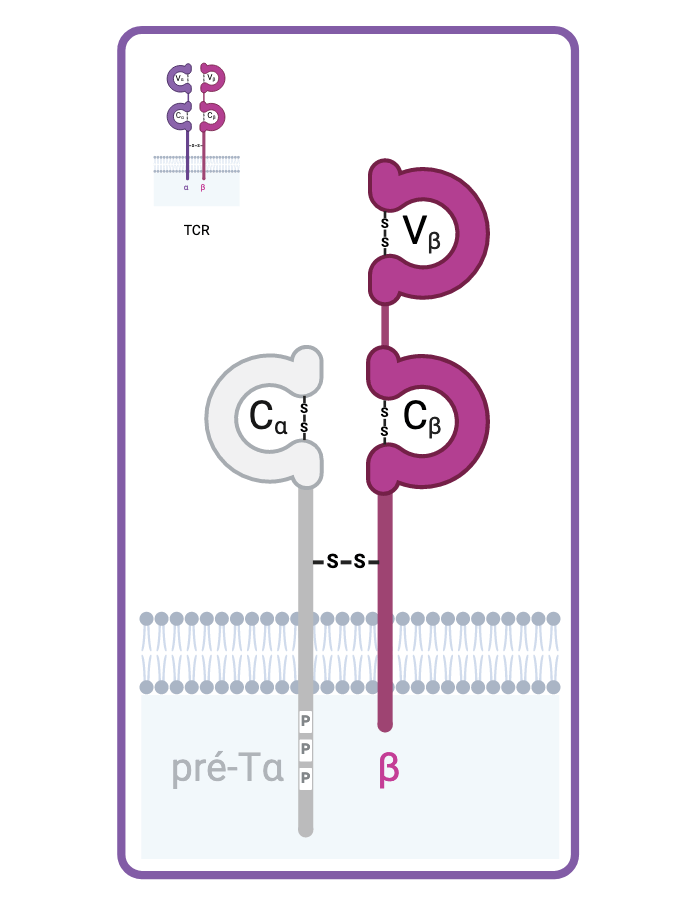
Si le réarrangement VDJ de la chaine β est non-productif, alors le pré-TCR n’est pas fonctionnel et le réarrangement de la chaine α ne peut pas débuter. C’est la β-séléction.
Si le réarrangement VDJ de la chaine β est productif, alors le pré-TCR est fonctionnel et son engagement induit une cascade d’événements notamment :
- L’arrêt du réarrangement de la chaine β
- Une prolifération importante
- L’initiation du réarrangement d’une chaine α
L’association d’une chaine β avec une chaine α aboutit à la synthèse du TCR.
Conséquences
- La β-sélection limite la synthèse de clones qui seraient inutiles en raison d’une chaine β non-fonctionnelle qui conduirait à un TCR non fonctionnel.
- L’arrêt du réarrangement de la chaine β induit l’exclusion allélique pour la chaine β.
- La concomitance de la prolifération importante avec l’initiation du réarrangement d’une chaine α a pour conséquence l’augmentation de la diversité potentielle du TCR, car pour une chaine β synthétisée, de multiples clones avec cette même chaine β et différentes chaines α vont pouvoir apparaitre.
L’acquisition du TCR est totalement aléatoire, indépendante des molécules HLA de l’individu comme de l’ensemble des antigènes, qu’ils soient du « soi » ou non. Il est donc nécessaire que l’ontogenèse des lymphocytes T comporte des étapes de sélection pour éliminer les clones inutiles d’une part et auto-réactifs d’autre part.
Étapes de sélection clonale
Une fois le TCR généré, deux étapes de sélection se succèdent :
La sélection corticale positive
Dans le cortex thymique, au contact des cellules épithéliales thymiques corticales (cTEC) exprimant un grand nombre de molécules HLA de classe I et de classe II.
Au stade précoce du développement du thymocyte, lorsqu’il est double négatif (CD4- CD8-)
Les cTEC expriment les antigènes du soi à la fois via les molécules HLA de classe I (présentation antigénique standard de toute cellule nucléée) ou via les molécules HLA de classe II grâce aux autophagosomes. Les phagosomes ayant la capacité de fusionner avec l’endosome où se trouvent les molécules HLA de classe II, cela permet aux cTEC de présenter des peptides intra-cellulaires – ceux qu’elles réabsorbent via les autophagosomes - dans les molécules HLA de classe II. (voir présentation antigénique)
Les lymphocytes doubles positifs ont trois destinées possibles en fonction de l’affinité de leur TCR pour les molécules HLA de classe I et/ou II exprimées par les cellules épithéliales thymiques corticales :
- Le TCR ne reconnait pas les molécules HLA du soi (90-95% des cas) : ces lymphocytes sont inutiles et entrent en apoptose par défaut de signal de survie (mort par négligence).
- Le TCR reconnait les molécules HLA du soi avec une trop forte affinité (2-5% des cas) : ces lymphocytes ont un potentiel auto-réactif élevé qui menace l’intégrité du soi et dégénèrent également.
- Le TCR reconnait les molécules HLA du soi avec une affinité modérée (2-5% des cas) : ces lymphocytes reçoivent des signaux de survie.
Conséquences ?
Ces étapes de sélection corticale permettent principalement de maintenir les clones lymphocytaires dont l’affinité pour les molécules HLA du soi est modérée. Les clones subsistant après l’étape de sélection positive deviennent simple positifs, CD4+ ou CD8.
On utilise aussi le terme de restriction au CMH pour parler de la sélection positive, car cette sélection est basée sur la reconnaissance des molécules HLA par le TCR.
Le HLA est le Complexe Majeur d’Histocompatibilité (CMH) de l’Homme.
La sélection médullaire négative
Où ?
Dans la médulla thymique, au contact des cellules épithéliales thymiques médullaires (mTEC) exprimant un grand nombre de molécules HLA de classe I et de classe II.
Quand ?
Suite à la sélection positive, les thymocytes sont alors simple positifs (CD4+ ou CD8+).
Mécanisme ?
Les cellules épithéliales thymiques médullaires synthétisent la protéine AIRE (AutoImmune REgulator) qui permet l’expression ectopique d’antigènes tissulaires. Cela permet qu’un maximum d’antigènes du soi puissent être exprimés par les mTEC.
La présentation des peptides du soi se fait à la fois :
- par les cellules épithéliales thymiques médullaires :
Elles expriment les antigènes du soi à la fois via les molécules HLA de classe I (présentation antigénique standard de toute cellule nucléée) ou via les molécules HLA de classe II grâce aux autophagosomes. Les phagosomes ayant la capacité de fusionner avec l’endosome où se trouvent les molécules HLA de classe II, cela permet aux cellules épithéliales thymiques médullaires de présenter des peptides intra-cellulaires – ceux qu’elles synthétisent de façon ectopique - dans les molécules HLA de classe II (voir présentation antigénique).
- par les cellules dendritiques présentes dans la médulla thymique :
Après qu’elles ont ingéré les antigènes ectopiques synthétisés par les cellules épithéliales thymiques médullaires, soit parce que la mTEC est en apoptose soit parce qu’elle secrète ces antigènes, les cellules dendritiques présentent ces antigènes via la classe I et la classe II (voir présentation antigénique).
Les cellules dendritiques et les macrophages présentent aussi de façon tolérogène des antigènes du non-soi auxquels l’organisme est fréquemment exposé comme ceux provenant des différentes flores commensales ou de l’ « environnement ordinaire ».
Conséquences ?
La sélection négative qui survient dans la médulla thymique permet de supprimer les clones qui reconnaissent les antigènes du soi et les antigènes exogènes fréquemment rencontrés avec une trop forte affinité. Par conséquent, elle permet d’induire une tolérance au soi et aux antigènes exogènes fréquemment rencontrés, comme ceux appartenant aux flores commensales ou à l’ « environnement ordinaire ».
Activation spécifique suite à la rencontre antigénique
L’activation des lymphocyte T est décrite dans la page mécanisme « activation des lymphocytes T ».